
Bakteriofág

Ahogy bakteriofágok vagy egyszerűen fágok ( egyes szám fág , az , az ókori görög βακτήριον baktērion , pálcika „és φαγεῖν phagein , enni”) kifejezés a különböző csoportok vírusok , hogy a baktériumok a gazdasejt szakosodott. A gazdaszervezet sajátossága szerint a fágokat taxonómiai csoportokra osztják , például coli , staphylococcus , diftéria vagy Salmonella bakteriofágokra. A becslések szerint több 10 30 virionok minden tengervíz, fágok sokkal gyakoribbak, mint bármilyen típusú a élőlénynek és a forma az úgynevezett virioplankton , köztük sok cyanophages (bakteriofágjának cianobaktériumok ).
Hagyományosan az archaea vírusait ( archaeovírusok, archealis vírusok, archaeovírusok ) néha még mindig (bakterio) fágoknak nevezik, ami még mindig az 1970-es évek elejére emlékeztet, amikor az archaea még nem volt megkülönböztetve a baktériumoktól és a fejfágú fágoktól . kutattak ( Caudoviricetes ), amelyek részben baktériumokat, részben archeákat fertőznek.
Kérjük, vegye figyelembe: A vírusok nem élőlények, mert nincs saját anyagcseréjük . Néhány tudós "életközeli" -nek nevezi őket.
sztori
A fágok hatását először 1917-ben írta le a francia-kanadai Félix Hubert d'Hérelle . Bár az angol Frederick Twort két évvel korábban már megfigyelte a bomlási folyamatokat a staphylococcus kultúrákban, amelyek a bakteriofágok hatására vezethetők vissza, publikációját gyakorlatilag figyelmen kívül hagyták. D'Hérelle tehát a bakteriofágok egyik felfedezője, az úgynevezett "baktériumfogyasztók" Frederick Twort mellett. Nevüket és felfedezésüket azonban d'Hérelle -nek köszönhetik. A d'Hérelle -vel párhuzamosan Philalethes Kuhn német mikrobiológus a bakteriális paraziták létezését feltételezte a bakteriális kultúrákban bizonyos körülmények között bekövetkezett változások megfigyelései alapján. Ezt Pettenkoferien néven említette, és a d'Hérelle által leírt „láthatatlan mikrobát, amely ellensúlyozza a vérhasot” e paraziták különleges eseteként látta. Mint később kiderült, megfigyelései nem a bakteriális parazita létezésén alapultak, hanem csak az általa vizsgált baktériumok alakjának megváltozásán.
D'Hérelle úgy képzelte el a bakteriofágot, mint egy "láthatatlan, korpuszkuláris élőlényt", amely alapvető formában létezik, és alkalmazkodik a különböző gazdaszervezetekhez, azaz a baktériumokhoz. Valójában, amennyire ma tudjuk, a bakteriofágok magasan specializált vírusok, amelyek egy adott gazdaszervezethez kötődnek. Még ha házigazdákról is beszélünk ebben az összefüggésben, a mai definíció szerint a bakteriofágok nem paraziták , mivel nem élőlények, mint vírusok . Az első vizsgált fág hét Escherichia coli baktériumból származó fág volt . Max Delbrück nevezte őket felfedezésük sorrendjében, mint Type ( English Type ) 1 (T1), Type 2 ( T2 ) és így tovább. Ezeknek a fágtörzseknek a jelenlegi rendszertani besorolása az ICTV szerint fajok és családok szerint a következő:
- T1: Escherichia T1 faj , Drexlerviridae
- T2: Escherichia vírus T4 (más néven T-páros fágok ), Myoviridae
- T3: Escherichia T3 faj , Autographiviridae
- T4: Az Escherichia vírus T4 típusú törzse .
- T5: Escherichia T5 faj , Demerecviridae
- T6: az Escherichia vírus T4 faja is
- T7: Escherichia vírus T7 , Autographiviridae
Ezt a hét fágot néha a T fágok (en. T fágok ) gyűjtőnév alá csoportosítják , de ez nem jelent családi csoportot ( taxont ). Ehelyett ezeket a bakteriofágokat néhány váltás után az ICTV fent megadott családokhoz rendeli (2021. januárjától). Csak a páros típusszámú fágok ( T-páros fágok ) bizonyultak szoros rokonságnak véletlenül, így egy fajt az ICTV taxonjaként hoztak létre. A páratlan számú típusok ( T-páratlan / T-páratlan fágok ) nem képeznek taxont. Mindezek a fágtípusok azonban fej-farok struktúrát alkotnak együtt, amit a vírusrend Caudovirales jellemzőinek képviselőiként látnak . Később ezt a gyakorlatot részben más szerzők is folytatták más caudovírusok elnevezésekor (pl. " T12 ", javaslat, családi megbízatás nélkül).
Építkezés


2 farok
3 nukleinsav
4 kapszid
5 gallér
6 hüvely
7 farokrost
8 tüske
9 alaplemez
A fej-farok felépítésű bakteriofágok alakját ( Caudovirales rend ) elsősorban az Escherichia coli T sorozat (T sorozat) fágjain derítették ki . A T2 bakteriofág egy 100 nm hosszúságú poliéderes fejből áll, amelynek farka megközelítőleg azonos hosszúságú. A bakteriofágokat csoportosítva taxonómiailag aszerint, hogy azok morfológiája , genom és fogadó . Különbséget tesznek az egyszálú DNS- sel rendelkező DNS- fágok, az úgynevezett ssDNA-fágok (az angol egyszálúak ) és a kettős szálú DNS-ek, az úgynevezett dsDNS-fágok között (angol kettős szálúak ). Az itt példaként kezelt T -sorozat Escherichia coli fágjai az utóbbi csoportba tartoznak.
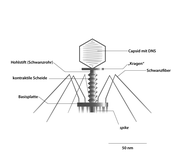
A már fentebb említett T-fágokat (például a Tequattrovirus nemzetséget Escherichia vírus T4 fajjal ), a Caudovirales rend többi tagjával együtt, viszonylag összetett, „fej-farok szerkezetű” szerkezet különbözteti meg a többi bakteriofágtól: egy alaplemezből (9), egy injekciós eszközből (injekciós készülék vagy farok, 2) és egy fejből (1) áll, amelyek az úgynevezett kapszidból (4) és a benne lévő nukleinsavból állnak ( genom , 3). A fej és az injektor / farok modul nyakkal van összekötve (gallér, 5). Az alaplemezt (amely a kapszidhoz és az injekciós készülékhez hasonlóan fehérjékből áll ) farokrostok (7) és tüskék (8) borítják, amelyeket a gazdasejt falán történő adszorpcióra használnak . Az injekciós készülék egy vékony csőből (farokcső, 6) áll, amelyen keresztül a fág nukleinsavat (3) injektálják a gazdasejtbe. A csövet összehúzódó farokhüvelybe csomagolják, amely összehúzódik az injekció beadása során. A kapszid 152 kapszomerből áll, ikozaéder szimmetriával, és tartalmazza a fág DNS -ét. Ennek a szerkezetnek köszönhetően a Tquattrovirus nemzetség fágjai ( Myoviridae család ) a szerkezetileg legösszetettebb vírusok közé tartoznak.

Az egyszálú DNS-t tartalmazó fágok általában kicsi, gömb alakúak és farok nélküli vagy fonalasak. A szintén előforduló RNS-fágok általában (ha eddig le vannak írva) egy fehérjeburokból állnak, amely egyszálú RNS-molekulát foglal magában. Ezen fágok átmérője körülbelül 25 nm, így az egyik legkisebb fág.
- Különböző fágok
T2 fág ; Axiálisan vágott kapszid

Lysehof a Bacillus fág gamma a Bacillus anthracis , a jobb fertőzött egyetlen telepet

Synechococcus S-PM2 fág tengervízből

Lambda fág ( Escherichia vírus lambda ); sematikus keresztmetszete




Szorzás

Saját anyagcsere hiányában a vírusoknak gazdaszervezetre van szükségük a szaporodáshoz , bakteriofágok esetén megfelelő, élő baktériumsejtre. A reprodukció öt fázisra osztható:
- Adszorpció specifikus sejtfal receptorokon : Az adszorpció során a farokrostok végei a baktérium felszínén lévő megfelelő molekulákhoz ( receptorokhoz ) kapcsolódnak.
- A fág nukleinsav injektálása a gazdasejtbe: A fág saját nukleinsava , a DNS vagy az RNS bejut a baktériumba. Az üres fágburok immár működésképtelen fehérjéi kívül maradnak a baktérium felszínén.
- Várakozási fázis: Ebben a fázisban nem lehet fágokat kimutatni a baktériumban. A transzkripciójának a virális genom , a fordítását a virális mRNS és a replikációját a virális nukleinsav most kezdődik . Ez a folyamat legfeljebb néhány órát vesz igénybe.
- Termelési fázis : Miután a fággének rögzített sorrendben aktiválódtak, minden víruskomponens, burokfehérje és farokrost kialakul.
- Érettségi fázis: A morfogenezis ezen fázisában az összeszerelés úgy történik , hogy érett fágrészecskéket képezzen. Először egy fejrészt, a kapszidot alakítják ki. A benne lévő fehérjék helyőrzőként szolgálnak, és később a kapszidba behatoló fág nukleinsav váltja fel őket. A nukleinsavszálak helytakarékos formát öltenek, mint egy gyapjúgolyó.
- Kibocsátás: A kész vírusrészecskék a gazdasejt enzimatikus oldódásával szabadulnak fel . A lizozim , amelyet az átprogramozott baktérium hozott létre, feloldja a bakteriális egér sejtfalat . A sejt felrobban, és körülbelül 200 fertőző fág szabadul fel.
Bizonyos típusú fágok esetében a reprodukció nem mindig a fent leírt lítikus séma szerint megy végbe . Abban az esetben, mérsékelt fágok különbséget kell tenni a lizogénaktivitását és litikus reprodukciós ciklus vagy fertőzés ciklus. Egy lizogén ciklusban a fág DNS -e beépül a baktérium kromoszómájába , és így létrejön egy próféta . Minden további sejtosztódás során a fág és a baktérium génjei megismétlődnek és együtt továbbadódnak. Ez a ciklus később a lítikus ciklushoz vezethet.
Óriási fágok
.jpg)
Az 540 kbp- nél nagyobb genomméretű kétszálú DNS-fágokat megafágoknak, míg a 200 kbp-nál nagyobb méretű fágokat jumbo-fágoknak nevezzük. 2018/2019 -ben a szerzők Bangladesben és Tanzániában élő emberek ürülékét , valamint afrikai páviánokat és Dániában sertéseket vizsgáltak. A minták a Prevotella ( Prevotellaceae ) nemzetség baktériumait tartalmazták , amelyeket számos dsDNS megafág fertőzött meg, amelyeket a szerzők „ Lak -fágoknak” neveztek el ( Laksam Upazila , Banglades) alapján. A talált fágokat (ideiglenesen) Lak-A1, Lak-A2, Lak-B1-Lak-B9 és Lak-C1 jelöléssel látták el. Laza filogenetikai kapcsolat lehet a "Sphingomonas Phage PAU" -val (ez az óriási fág megfertőzi a Sphingomonas paucimobilis , Sphingomonadaceae faj baktériumait ) és így a Myoviridae fágcsaláddal . A szerzők arra a következtetésre jutnak, hogy a "lac -fágok" "széles körben elterjedtek, de korábban figyelmen kívül hagyták a bél mikrobióma tagjait".
2020 februárjában Basem Al-Shayeb és munkatársai elemzést tettek közzé, amely folytatja ezeket a vizsgálatokat. Ebben 500 kbp-n húzzák meg a megafágok határát (ami nyilvánvalóan bázispárokat jelent a kétszálú esetben és bázisokat vagy nukleotidokat az egyszálú esetben). A szerzők szívesebben tekintenek minden 200 kbp -nál nagyobb fágot (azaz jumbo -fágot és megafágot) " angol hatalmas fágoknak " (itt óriási fágoknak fordítják). A szerzők tíz csoportot azonosítottak ebben a csoportban , amelyekhez a következő neveket javasolták: "Kabirphage", "Mahaphage" (beleértve a " Lak -fágok csoportját "), "Biggiephage", "Dakhmphage", "Kyodaiphage" , "Kaempephage", "Jabbarphage", "Enormephage", "Judaphage" és "Whopperphage" (minden név a "hatalmas" vagy " hatalmas " kifejezésre utal a szerzők különböző nyelvein). Különböző minták metagenomikus elemzésén keresztül 351 dsDNS fágszekvenciát tudtak azonosítani, amelyek közül csak 3 volt kevesebb, mint 200 kbp. A legnagyobb genom 735 kbp hosszú volt (Mahaphage, ami láthatóan új rekord; az előző 596 kbp); a közönséges, nem óriási fágok átlaga mindössze 52 kbp. Úgy tűnik, hogy néhány óriási fág nem szabványos genetikai kódot használ, amelyben az UAG stopkodon egy aminosavat kódol . A házigazdák (többnyire) a Firmicutes vagy a Proteobacteria baktériumai , de - mint a "Lak -fágokkal " rendelkező Mahaphage csoport tagjai esetében - a Bacteroidetes is . A genom kódolja a tRNS -eket a fágokban általánosan használt fehérjék mellett . A fágok kölcsönhatásba lépnek a CRISPR / Cas rendszerben is (lásd CRISPR , CRISPR / Cas módszer , genomszerkesztés ): a rendszer minden fő típusa képviseltette magát, de úgy tűnt, hogy a legtöbb fág a gazdaszervezet Cas -fehérjéit használja fel a védekezéshez. Ezenkívül úgy tűnt, hogy a fágok segítik a házigazdák CRISPR immunrendszerét a versengő fágok elleni küzdelemben. Néhány Pseudomonas -fertőző fág az anti-CRISPR-eket (Acrs) és a fehérjéket is kódolja, amelyek egy maghoz hasonló rekeszt képeznek, amelyben a fág a gazdától függetlenül jobban meg tudja replikálni genomját (lásd Viroplasma ). A szerzők munkájukban további bizonyítéknak tekintik az óriási fágok világméretű elterjedését. Bizonyítékot találtak arra, hogy a fágok különböző gazdaszervezetek és ökoszisztémák között vándoroltak, ami hatással van a toxin- és antibiotikum -rezisztencia gének terjedésére . A CRISPR eszközeit a jövőben fel lehet használni a CRISPR / Cas "génolló" fejlesztésére és funkcionalitásának bővítésére.
Egy másik óriási fág az A9 Megasphaera fág (más néven Huge Phage A9 ), nem tévesztendő össze az A9 Brochothrix fággal ( Brochothrix vírus A9 , Herelleviridae faj ).
Farok nélküli fágok
A kutatás sokáig csak a Caudovirales rend tagjait vette figyelembe , amelyek képviselői fágok (bakteriális és régészeti vírusok), fej-farok szerkezettel. Csak a közelmúltban váltak a "fark nélküli" fágok a kutatás tárgyává. Néhány képviselő:
- Rendeljen Tubulavirales -t (fonalas bakteriofágokat) és másokat. családi Inoviridae
- Finnlakeviridae család (ssDNS, Flavobacterium vírus FLiP fajokkal, más néven Phage FLiP )
- " Autolykiviridae " család (dsDNS, javasolt)
- Faj Planktothrix-Phage PaV-LD ( Cyanophage PaV-LD )
crAssphages és Gubaphages
Camarillo-Guerrero, Almeida és mtsai. A 2019/2020 leírja az emberi bélflóra bakteriofágokkal kapcsolatos metagenomikus elemzésének eredményeit . Azt, hogy ez egy új clade, az úgynevezett "Gubaphagen" ( angolul G ut Ba cteroidales fág , Gubaphage kládja ) (két nemzetségek, G1 - fertőzött Bacteroides és G2 - fertőzött Parabacteroides [s] ), amely szerint a durva fág körülbelül 10 A nemzetségek ( crAsslike fágok , a Caudovirales rend eredetileg javasolt tagjai , Podoviridae család , körülbelül 10 nemzetséggel) képviselik a második leggyakoribb vírust (azaz bakteriofágot) ebben a környezetben. A Gubaphages jellemzői a "p-crAssphage" -ra emlékeztetnek.
Időközben felmerült az a javaslat, hogy a crAssphages saját rendjét adja meg a Crassviralesnek a Caudoviricetes osztályon belül . A crAssphageshez való hasonlóságuk miatt a Gubaphages valószínűleg a Crassvirales (vagy legalábbis a Caudoviricetes ) tagjai is lennének .
Halovírusok
A halofil baktériumokat és archeákat megtámadó fágokat informális (nem taxonómiai) halovírusok ( angol halovírusok ) néven osztályozzák . Az ICTV által megerősített Halobacterium vírus phiH fajtájú Myohalovirus ( Caudovirales : Myoviridae ) nemzetségen és a javasolt "Halorubrum fág HF2" fajon kívül ezek még besorolás nélküli és még nem megerősített "HF1", "HCTV-1" fajok , "2" és "5", "HGTV-1", "HHTV-1" és "2", "HRTV-4", "5", "7" és "8" ( caudovirales ), "HSTV-1" "( caudovirales : Podoviridae ) és" 2 "( Caudovirales : Myoviridae )," HVTV-1 "( Caudovirales : Siphoviridae )," Halovirus VNH-1 "(" VNH-1 ", Fuselloviridae ) és" Haloferax farok vírus 1 "( HFTV1, Caudovirales ).
Magroviruses
Az Euryarchaeota tengeri archeái a Marine Group ( English Marine Group ) II (MG-II, MG-IIa – MG-IId), III (MG-III) és IV (MG-IV) osztályba sorolhatók -a tengeri Az I. csoport (MG-I) a Thaumarchaeota tengeri archeáját jelöli .
A nem taxonómiai Magroviruses ( angolul magroviruses , MArine GROup II vírusok ) megjelölést az elsőként említett MG-II csoport Euryarchaeotájában élősködő fágok osztályozására használják. Ezek a dsDNS vírusok, amelyek genom mérete 65-100 kbp , fej-farok szerkezettel: "Magrovirus A", "Magrovirus B1" és "B2", valamint "Magrovirus C" és (feltételezett) "Magrovirus D". ".
alkalmazási területek
A fágok széles körű alkalmazást találtak az orvostudományban , a biológiában , az agrártudományokban , különösen a géntechnológia területén . Gazdaspecifikusságuk miatt a fágokat használják a gyógyászatban a bakteriális kórokozók meghatározására . Ezt az eljárást lizotípusnak nevezik . A többszörös antibiotikum -rezisztencia növekvő előfordulása miatt jelenleg intenzív kutatások folynak a bakteriofágok antibiotikum -helyettesítőként való felhasználásáról a humán gyógyászatban (lásd: fágterápia ). Problémák itt a fágok testben tapasztalható gyenge stabilitása miatt merülnek fel, mivel a fagociták nagyon rövid idő alatt eltávolítják őket idegen testként. Felix d'Hérelle (lásd fent) már jóval a penicillin és az antibiotikum felfedezése előtt felfedezte ezt a fágok alkalmazását bakteriális fertőzések kezelésére . Később azonban az antibiotikumokat alkalmazó kemoterápia bevezetésével a fágterápiát kivitelezhetetlennek tartották, és elfelejtették. D'Hérelle 1934 -ben Georgi Eliava grúz mikrobiológussal együtt megalapította az Eliava Fágkutató Intézetet a Grúz Szovjet Szocialista Köztársaságban , amely ma is létezik. Ma az egyébként terápia-rezisztens bakteriális fertőzések fágterápiáját ott és a wroclawi Ludwik Hirszfeld Immunológiai és Kísérleti Terápiás Intézetben végzik (a Lengyel Tudományos Akadémia része ) . Németországban a terápiás célú felhasználás még nem engedélyezett.
Az élelmiszertermelésben való alkalmazás sokrétű; Például kolbászcsomagoláskor vagy sajtdaraboláskor fág spray -t használnak.
A géntechnológia , mérsékelt fágokat használjuk a vektorok (például fág λ ). Ebből a célból a fágokat úgy állítják elő, hogy a virulenciát okozó géneket kivessék a genomjukból, és olyan génekkel helyettesítsék, amelyek géntechnológiai célokra érdekesek, például az inzulin termeléséhez szükséges génekkel . Ezeket a módosított fágokat ezután érintkezésbe hozzák megfelelő baktériumokkal, például E. colival . Miután ellenőrizte, hogy a kívánt gént beépítették-e a bakteriális genom genetikai anyagába (az egyik olyan gén-expresszált antibiotikum-rezisztenciát használ, amely a kívánt klónozandó génekhez kapcsolódik ), a módosított baktériumsejtek tovább tenyészthetők és az inzulin előállítható ez az eset elkülöníthető. Hasonlóképpen, fágok használnak mezőgazdasági technológia a transzdukálhatósága bizonyos gének a növények . Egy fontos alkalmazás biokémiai a fág display azonosításához kötőpartnerek, például B. az új hatóanyagok izolálásában.
Azonban könnyebb, mint a fágok segítségével átalakítani a szabad DNS -t, amelyet manapság főleg bakteriális sejtekbe történő átvitelre használnak.
A fágokat és összetevőket az élelmiszerekben található mikrobiális szennyeződések (pl. Affinitás mágneses elválasztás ), valamint az endotoxinokkal szennyezett laboratóriumi minták eltávolítására használják . Ezenkívül léteznek humán diagnosztikai alkalmazások, különösen a kórokozó kórházi baktériumok, például az MRSA dekolonizációjának klinikai területén . A fág fehérjék lehet optimalizálni az adott alkalmazás útján fehérje design.
Lehetséges gazdasági károk
A bakteriofágok kárt okozhatnak mindenütt, ahol a bakteriális folyamatok szolgálják az embereket, és kívánatosak. A tejsavbaktériumok (LAB) fertőzése nyers tejből származó fágokkal a leggyakoribb oka a csökkent vagy hiányzó enzimaktivitásnak a sajt- vagy túrótej -termelő induló kultúrákban .
osztályozás
A prokarióta vírusok (bakteriális és archeális vírusok, "bakteriofágok") nem alkotnak zárt családi csoportot ( taxont ). Sok fágcsoport esetében még mindig vannak informális nevek a gazdájuk után (lásd fent), pl. B. Cyanophages ( cianobaktériumok ), Coliphages (Colibacterium E. coli ), ezek szintén nem kapcsolódó csoportok. Másik különlegessége a műholdas vírusok , amelyeknek helper vírusok bakteriofágok, ezek néha műholdas fágok. Az egyik példa az "Escherichia Phag P4", amely segítő vírusként a P2 ( Myoviridae , Peduovirus nemzetség) kolifágot igényli .
Baltimore -i besorolás
A Baltimore -féle osztályozás szerint a fágokat genomjuk szerkezete szerint az alábbiak szerint csoportosíthatjuk:
- dsDNS bakteriofágok:
- Család: Ackermannviridae
- Család: Autographiviridae
- Család: Chaseviridae
- Család: Demerecviridae
- Család: Drexlerviridae
- Család: Herelleviridae
- Család: Myoviridae
- Család: Podoviridae
- Család: Siphoviridae
- Család: Tectiviridae
- Család: Corticoviridae
- Család: Plasmaviridae
- Család: Lipothrixviridae
- Család: Rudiviridae
- Család: Fuselloviridae
- Család: Halspiviridae (nemzetség: Salterprovirus és Salterprovirus His1 alias His 1 vírus )
- Család: Guttaviridae
- Család: Bicaudaviridae
- Család Thaspiviridae (a nemzetség Nitmarvirus és fajok Nitmarvirus NSV1 alias Nitrosopumilus maritimus vírus 1 )
- "Autolykiviridae" család (javasolt, lásd fent)
- ssDNS bakteriofágok:
- Finnlakeviridae család
- Család: Inoviridae - javasolt felosztás metagenóm adatok alapján: "Amplinoviridae", "Protoinoviridae", "Photinoviridae", "Vespertilinoviridae", "Densinoviridae" és "Paulinoviridae"
- Család: Plectroviridae
- Család: Microviridae
- Különleges eset
- Család Pleolipoviridae (a nemzetség Gammapleolipovirus és fajok Gammapleolipovirus His2 alias His 2 vírus , haloarcula vírus His2 )
- dsRNS bakteriofágok:
- Család: Cystoviridae
- ssRNS bakteriofágok:
- Család: Fiersviridae (más néven Leviviridae )
Taxonómiai besorolás az ICTV szerint
A vírusok rendszertani rendszerében , a Nemzetközi Vírusbiztonsági Bizottság (ICTV) szerint , a fágok a következő rendszertani csoportokban találhatók:
| terület | rendelés | család | morfológia | Genom | Példák |
|---|---|---|---|---|---|
| Riboviria | Levivirales | Leviviridae | un burkolta , izometrikus | ssRNS , lineáris | MS2 , Qβ |
| Mindivirales | Cystoviridae | burkolt, gömb alakú | dsRNS , szegmentált | Phi6 | |
| Varidnaviria | Belfryvirales | Turriviridae | burkolt, izometrikus | dsDNS , lineáris | STIV1 |
| Halopanivirales | Sphaerolipoviridae | burkolt, izometrikus | dsDNS, lineáris | SH1 fág | |
| Kalamavirales | Tectiviridae | fedetlen, izometrikus | dsDNS, lineáris | PRD1 | |
| Vinavirales | Corticoviridae | fedetlen, izometrikus | dsDNS, kör alakú | PM2 | |
| Duplodnaviria | Caudovirales | Ackermannviridae | fedetlen, összehúzódó farok | dsDNS, lineáris | ϕMAM1 |
| Autographiviridae | fedetlen, összehúzódó farok | dsDNS, lineáris | Acintetobacter fág P2 | ||
| Myoviridae | fedetlen, összehúzódó farok | dsDNS, lineáris | T4 , Mu , P1 , Coliphage P2 | ||
| Siphoviridae | fedetlen, nem összehúzódó farok (hosszú) | dsDNS, lineáris | λ , T5 , HK97 , N15 | ||
| Podoviridae | fedetlen, nem összehúzódó farok (rövid) | dsDNS, lineáris | T7 , T3 , Φ29 , P22 | ||
| Monodnaviria | Haloruvirales | Pleolipoviridae | burkolt, pleomorf | ssDNA , cirkuláris / dsDNS, cirkuláris / dsDNS lineáris | HHPV1 , HRPV1 |
| Petitvirális | Mikrovírusok | fedetlen, izometrikus | ssDNS, kör alakú | X174 | |
| Tubulavirales | Inoviridae | fedetlen, fonalas | ssDNS, (többnyire) kör alakú | M13 , CTXφ | |
| nem kijelölt | Vírusos kötés | Lipothrixviridae | burkolt, rúd alakú | dsDNS, lineáris | AFV1 |
| Rudiviridae | fedetlen, rúd alakú | dsDNS, lineáris | SIRV1 | ||
| nem kijelölt | nem kijelölt | Ampullaviridae | becsomagolva, palack alakú | dsDNS, lineáris | ABV |
| Bicaudaviridae | fedetlen, citrom alakú | dsDNS, kör alakú | ATV | ||
| Clavaviridae | fedetlen, rúd alakú | dsDNS, kör alakú | APBV1 | ||
| Finnlakeviridae | dsDNS | FLiP | |||
| Fuselloviridae | fedetlen, citrom alakú | dsDNS, kör alakú | SSV1 | ||
| Globuloviridae | burkolt, izometrikus | dsDNS, lineáris | PSV | ||
| Guttaviridae | fedetlen, tojásdad | dsDNS, kör alakú | SNDV , APOV1 | ||
| Plasmaviridae | burkolt, pleomorf | dsDNS, kör alakú | L2 fág | ||
| Portogloboviridae | burkolt, izometrikus | dsDNS, kör alakú | |||
| Spiraviridae | fedetlen, rúd alakú | ssDNS, kör alakú | ACV | ||
| Tristromaviridae | burkolt, rúd alakú | dsDNS, lineáris | TTSV1 |
A Picobirnaviridae család tagjai ( Durnavirales rend ) szintén a baktériumokat fertőzik, nem az emlősöket.
Egy másik javasolt fágcsalád az „ Autolykiviridae ” (dsDNA).
irodalom
- Nicholas H. Mann: A fágok harmadik kora. In: PLOS Biology . 3. kötet, 5. szám, 2005. május 17., e182. Cikk, doi: 10.1371 / journal.pbio.0030182 ( teljes szöveg PDF -ként ).
- Nancy Trun, Janine Trempy: Bakteriofág. In: Nancy Jo Trun, JE Trempy, Janine Trempy: Alapvető bakteriális genetika. Blackwell, Oxford 2003, ISBN 0-632-04448-9 ( teljes szöveg PDF formátumban ).
- Górski A, Weber-Dabrowska B: Az endogén bakteriofágok lehetséges szerepe a betolakodó kórokozók ellenőrzésében . In: Cellular and Molecular Life Sciences . 62. évfolyam, 5. szám, 2005. március, 511–519. doi : 10.1007 / s00018-004-4403-6 . PMID 15747058 .
- Forest Rohwer, Merry Youle, Heather Maughan, Nao Hisakawa, Leah L Pantéa: Élet a fágvilágunkban: centenáriumi terepkalauz a Föld legkülönfélébb lakóinak. Wholon, San Diega (CA) 2014, ISBN 978-0-9904943-0-0 .
- Hans Günther Schlegel, Georg Fuchs (szerk.): Általános mikrobiológia. 8. kiadás, Thieme, Stuttgart 2006, ISBN 3-13-444608-1 .
- Jong-Geol Kim, So-Jeong Kim, Virginija Cvirkaite-Krupovic, Mart Krupovic et al. : Az orsó alakú vírusok megfertőzik a tengeri ammóniát oxidáló thaumarchaeát. In: Proceedings of the National Academy of Sciences . (PNAS) 116. kötet, 31. szám, 2019. július, 201905682. Cikk, doi: 10.1073 / pnas.1905682116 ( teljes szöveg online ).
web Linkek
- Építs és szaporíts animációval
- Bakteriofágok és fágterápia: kérdések és válaszok áttekintése, a Braunschweig -i DSMZ (German Collection of Microorganisms and Cell Cultures GmbH) Leibniz Intézet információi
- Szövetségi Kockázatértékelési Intézet (BfR) : Kérdések és válaszok a bakteriofágokkal kapcsolatban
- Az Eliava Intézet honlapja
- Fágterápia a Ludwik Hirszfeld Immunológiai és Kísérleti Terápiás Intézetben, Breslau
- Fágterápia tüdőgyulladás ellen , kutatási projekt Párizsban
- Phagoburn , EU kutatási projekt égési sérülések fágterápiájáról
- "Baktériumfalók" antibiotikumok helyett , VDI-Nachrichten, 2015. szeptember 25
- Gyógyító vírusok - Fertőzések leküzdése a bakteriofágokkal Rádió adása, Bavaria 2, 2012. szeptember 29, hozzáférés: 2015. szeptember 25
- A bakteriális ragadozó segíthet csökkenteni a COVID -19 haláleseteket - „potenciális játékváltó”. Be: 2020. június 26, Forrás: Birminghami Egyetem
- Az antibiotikum -válságból kilépő vírusokkal. - Jean Crépu dokumentumfilmjeaz ARTE -ról (F 2019, 54 perc)
Egyéni bizonyíték
- ↑ NCBI: Bacillus fág Gamma (faj)
- ↑ a b Daniel Bojar: Hasznos baktériumölők. On: Spektrum.de - Spectrum of Science. 2020 júniusától, 40–45.
- ↑ SIB: Baktériumokat fertőző vírusok. Be: ViralZone .
- ↑ Mart Krupovic, Anja Spang, Simonetta Gribaldo, Patrick Forterre, Christa Schleper: A thaumarchaeal provirus tanúskodik a farkú vírusok ősi összefüggéséről az archaeával. In: Biokémiai Tranzakciók. 39. kötet, 1. szám, 2011. január, 82-88. O., Doi: 10.1042 / BST0390082 , PMID 21265751 .
- ^ F. d'Hérelle (1917): Sur un microbe invisible antagonist des bacilles dysentériques. In: CR Ac. Tudományok. 165. szám, 373-375.
- ^ Loos-Frank, Brigitte, Lane, Richard P.: A paraziták biológiája . 3., frissített és javított kiadás. Springer Verlag, Berlin 2018, ISBN 978-3-662-54862-2 , pp. 4 ( google.de [letöltve: 2019. március 17]).
- ↑ NCBI: T-fágok
- ↑ Rolf Sauermost, Doris Joyful és mtsai. : T fág. - Biológia lexikon. On: Spektrum.de , hozzáférve 2021. január 31 -én. A családi feladatok már nem felelnek meg az ICTV szerinti jelenlegi állapotnak.
- ↑ L. McKane, J. J. Ferretti: Fág -gazda kölcsönhatások és az A típusú streptococcus exotoxin termelése az A csoport streptococcusaiban . In: Fertőzés és immunitás . 34. kötet, 3. szám, 1981. december, 915–919. PMID 7037644 . PMC 350956 (ingyenes teljes szöveg).
- ↑ Victor Krylov, Maria Bourkaltseva, Elena Pleteneva, Olga Shaburova, Sergey Krylov, Alexander Karaulov, Sergey Zhavoronok, Oxana Svitich, Vitaly Zverev; Julie Thomas, Lindsay Black (szerk.): Phage phiKZ - Az első óriások. In: Vírusok. 13. kötet, 2. szám, Különkiadás Giant vagy Jumbo Phages. 149., 2021. január 20., doi: 10.3390 / v13020149 .
- ↑ a b Audra E. Devoto, Joanne M. Santini et al. : A megafágok megfertőzik a Prevotella -t, és változatai széles körben elterjedtek a bél mikrobiómákban. In: Természeti mikrobiológia. 4. kötet, 693-700. Oldal, 2019. január 28., doi: 10.1038 / s41564-018-0338-9 , különösen az 1. táblázat és a 11. kiegészítő ábra .
- ↑ Lakshminarayan M. Iyer, Vivek Anantharaman, Arunkumar Krishnan, A. Maxwell Burroughs, L. Aravind: Jumbo Phages: A Comparative Genomic Overview of Core Function and Adaptions for Biological Conflicts , in: MDPI Viruses, Volume 13, No. 1, Special Giant vagy Jumbo Phages, 2021. január 5., 63. szám; doi: 10.3390 / v13010063 , különösen kiegészítés (zip PDF és xlsx)
- ↑ NCBI: Sphingomonas fág PAU. (faj)
- ↑ Richard Allen White III, Curtis A. Suttle: A „Sphingomonas paucimobilis” HER1398 (Proteobacteria) törzs, a Giant PAU Phage házigazdája, a genomszekvencia -tervezet azt jelzi, hogy a „Sphingobacterium” (Bacteroidetes) nemzetség tagja. In: Genomközlemények. 1. kötet, 4. szám, 2013. július-augusztus, e00598-13, doi: 10.1128 / genomeA.00598-13 , PMID 23929486 , PMC 3738902 (ingyenes teljes szöveg)
- ↑ University of California - Berkeley: [1] , ScienceDaily, 2019. január 28.
- ↑ UCL: Új, óriási bakteriális vírus található az emberi bélben. University College London, 2019. január 29.
- ^ Colm Gorey: Az emberekben felfedezett óriási vírusok kérdéseket vetnek fel az életről. Bekapcsolva: siliconrepublic.com , 2019. január 29 -től.
- ↑ a b Basem Al-Shayeb, Rohan Sachdeva, L. Chen, Jillian F. Banfield et al. : Hatalmas fágok kládjai a Föld ökoszisztémáiból , itt: Természet 2020. február 12-től, doi: 10.1038 / s41586-020-2007-4 , bioRxiv : 10.1101 / 572362v1 ( előzetes teljes szöveg)
- ↑ Ed Yong: Hatalmas felfedezés a vírusok világában. Be: Az Atlanti -óceán. 2020. február 20 -tól.
- ↑ Michael Le Page: Óriási vírusok fegyveresítették fel a CRISPR -t bakteriális gazdájuk ellen. Be: NewScientist. 2019. március 30 -tól.
- ↑ Óriás bakteriofágok Hídrés az élő mikrobák és vírusos gépek között. Bekapcsolva: SCI-NEWS 2020. február 13 - tól.
- ↑ Tessa Koumoundouros: A tudósok óriási vírusokat fedeznek fel, amelyeknek az élő sejtekben csak korábban látott tulajdonságai vannak. Be: ScienceAlert 2020. február 14 -től.
- ↑ Daniela Albat: Rekord méretű genommal rendelkező fágot fedeztek fel: 2020. február 18, scinexx.de
- ↑ Jan Osterkamp: Az anti-CRISPR állítólag jobbá teszi a CRISPR-t. Bekapcsolva: Spektrum.de 2020. január 16 -tól.
- ↑ Annika Röcker: A genetikai olló tehetetlen néhány vírussal szemben. Bekapcsolva: Spektrum.de 2019. december 10 -től.
-
↑ Basem Al-Shayeb, Rohan Sachdeva, Lin-Xing Chen, Cindy J. Castelle, Alexander L. Jaffe, Jennifer A. Doudna , Jillian F. Banfield et al. : Hatalmas fágok lapjai a Föld ökoszisztémáiból. In: Természet . 578. szám, 2020. február 12., 425-431. Oldal, doi: 10.1038 / s41586-020-2007-4 . Továbbá:
- bioRxiv , PrePrint, 2019. március 11., doi: 10.1101 / 572362
- Hatalmas baktériumfogyasztó vírusok szüntetik meg a szakadékot az élet és a nem élet között-A nagy bakteriofágok bakteriális géneket hordoznak, beleértve a CRISPR-t és a riboszómás fehérjéket. On: eurekalert.org 2020 február 12 -től.
- ↑ Elina Laanto, Sari Mäntynen, Luigi De Colibus, Jenni Marjakangas et al.: A borealis tóban talált vírus összekapcsolja az ssDNA és a dsDNA vírusokat. In: Proceedings of the National Academy of Sciences. 114. kötet, 31. szám, 2017. július, doi: 10.1073 / pnas.1703834114 .
- ↑ a b Kathryn M. Kauffman, Fatima A. Hussain, Joy Yang és mtsai: A nem farkú dsDNS vírusok fő vonulata, mint a tengeri baktériumok felismerhetetlen gyilkosai. In: Természet. 554. kötet, 118-122. O., 2018. január 24., doi: 10.1038 / nature25474 .
- ^ A tudósok új típusú vírust találnak a világ óceánjaiban: Autolykiviridae. Be: sci-news 2018. január 25 - től.
- ↑ David L. Chandler: A kutatók felfedeznek egy hiányzó láncszemet a vírusfejlődésben. Be: SciTechDaily 2018. január 25 -től.
- ↑ A kutatók felfedeznek egy titokzatos vírust, amely uralja az óceánokat. Be: üzleti bennfentes 2018. január 29 -től.
- ↑ Soha nem látott, furcsa DNS-sel rendelkező vírusokat fedeztek fel az óceánban. On: sciencealert.com 2018. január 25 -től.
- ↑ NCBI: Autolykiviridae. (család) - osztályozatlan dsDNA vírusok.
- ↑ E-Bin Gao, Xiu-Ping Yuan, Ren-hui Li, Qi-Ya Zhang: Egy új, a fonalas cianobaktériumra „Planktothrix agardhii” (Cyanophyceae) fertőző cianofág izolálása a Donghu-tóból, Kínából. In: Vízi mikrobiológiai ökológia. (AME), 54. kötet, 1. szám, 2009. február, 163–170. O., Doi: 10.3354 / ame01266 ( teljes szöveg PDF formátumban ).
- ↑ Mélanie Gerphagnon, Deborah J. Macarthur, Delphine Latour, Claire M. M. Gachon, Floris Van Ogtrop, Frank H. Gleason, Télesphore Sime - Ngando: Mikrobiális játékosok, amelyek részt vesznek a fonalas és gyarmati cianobaktériumok virágzásában, a gombás parazitizmusra összpontosítva. In: sfam Environmental Microbiology. 17. kötet, 8. szám, 2573–2587. O., Doi: 10.1111 / 1462-2920.12860 ( teljes szöveg PDF formátumban ), különösen kiegészítés; Teljes szöveg PDF -ként 2. táblázat.
- ↑ NCBI: Planktothrix PAG-LD fág. (faj)
- ↑ CrAssphage: Korábban ismeretlen ősi bélvírus a fél világ lakosságában él. Be: sci-news 2014. augusztus 11-től ( angol ).
- ↑ Natalia Yutin, Kira S. Makarova, Ayal B. Gussow, Mart Krupovic, Anca Segall, Robert A. Edwards, Eugene V. Koonin: Egy kiterjedt bakteriofágcsalád felfedezése, amely magában foglalja az emberi bélből származó leggyakoribb vírusokat . In: Természeti mikrobiológia . 3, 2017. 1. szám, 38–46. doi : 10.1038 / s41564-017-0053-y . PMID 29133882 . PMC 5736458 (ingyenes teljes szöveg).
- ↑ Eugene V. Koonin: A papír mögött: A legelterjedtebb emberrel kapcsolatos vírus, amely már nem árva , 2017. november 13.
- ↑ SIB: crAsslike fágok. Be: viruszone.expasy.org - Rendelés: Caudovirales , Becsült körülbelül 10 nemzetség.
-
↑ Luis Fernando Camarillo-Guerrero, Alexandre Almeida, Guillermo Rangel-Pineros, Robert D. Finn, Trevor D. Lawley: Az emberi bél bakteriofág diverzitásának masszív bővülése , in: Cell Resource Volume 184, No. 4, pp. 1098-1109. e9 , 2021. február 18., doi: 10.1016 / j.cell.2021.01.029 . 2020. szeptember 3 -i előnyomtatás : bioRxiv , Európa PMC , doi: 10.1101 / 2020.09.03.280214 . Továbbá:
- Martin Vieweg: Több ezer típusú vírust fedeztek fel a bélflórában , On: Wissenschaft.de 2021. február 18 -tól (német)
- Daniel Lingenhöhl: Virobiom :
- A biológusok majdnem 143 000 bakteriofág fajt találnak az emberi bélben. Be: sci-news 2021. február 19 - től
- Peter Dockrill: A tudósok 140 000 vírusfajt találnak az emberi bélben , és a legtöbb ismeretlen. On: sciencealert.com 2021. február 28 -tól
- A tudósok több mint 140 000 vírusfajt azonosítanak az emberi bélben. Bekapcsolva: sciencedaily.com 2021. február 18 -tól
- ↑ Luis Fernando Camarillo Guerrero: Az emberi bélfág integratív elemzése metagenomikai módszerrel. PhD dolgozat, Gonville & Caius College, Cambridge -i Egyetem, 2020 augusztus, doi: 10.17863 / CAM.63973 .
- ^ Majd Turner, Andrew M. Kropinski, Evelien M. Adriaenssens: Útmutató a genom-alapú fágrendszerekhez . In: MDPI vírusok. 13. kötet, 3. szám, Bakteriális vírusok szakasz, 2021. március 18., 506. o., Doi: 10.3390 / v13030506 .
- ↑ Nina S. Atanasova, Hanna M. Oksanen, Dennis H. Bamford : Archaea, baktériumok és eukarióták halovírusai. In: Aktuális vélemény a mikrobiológiából. 2015. június 25., 40–48. O., Doi: 10.1016 / j.mib.2015.04.001 , PMID 25932531 .
- ↑ NCBI: Halovírusok (klád)
- ↑ NCBI: Myohalovírus (nemzetség)
- ^ ICTV: ICTV Taxonómia története: Halobacterium vírus phiH
- ↑ NCBI: Halorubrum fág HF2 (faj)
- ↑ a b c Yosuke Nishimura, Hiroyasu Watai, Takashi Honda et al.: A környezeti vírusgenomok új megvilágításba helyezik a vírus-gazda kölcsönhatásokat az óceánban. In: mSphere. 2. kötet, 2. szám, 2017. március-április, e00359-16, doi: 10.1128 / mSphere.00359-16 , PMC 5332604 (ingyenes teljes szöveg), PMID 28261669 , különösen a 4. ábra.
- ↑ a b c Darius Kazlauskas, Mart Krupovic, Česlovas Venclovas: A DNS-replikáció logikája a kétszálú DNS-vírusokban: a vírusgenomok globális elemzésének betekintése. In: Nukleinsavak kutatása. 44. kötet, 10. szám, 2016. június 2., 4551–4564. Oldal, doi: 10.1093 / nar / gkw322 , PMC 4889955 (ingyenes teljes szöveg), PMID 27112572 .
- ↑ a b c D Prangishvili, DH Bamford, P Forterre, J Iranzo, EV Koonin, M Krupovic: A rejtélyes archeális viroszféra . In: Nature Reviews Microbiology . 15. kötet, 12. szám, 2017. november 10., 724-739. doi : 10.1038 / nrmicro.2017.125 . PMID 29123227 . Lásd különösen az 1. ábrát
- ↑ NCBI: Halovírus HSTV-1 (faj)
- ↑ NCBI: Halovírus HSTV-2 (faj)
- ↑ NCBI: Halovírus HVTV-1 (faj)
- ↑ Anukriti Sharma, Matthias Schmidt, Bärbel Kiesel et al.: Bakteriális és archeális vírusok a Himalája meleg forrásaiból a Manikaran Modulate Host Genomes -ben. In: Határok a mikrobiológiában. 9. kötet, 2018. december 14., 3095. o., Doi: 10.3389 / fmicb.2018.03095 , PMC 6302217 (ingyenes teljes szöveg), PMID 30619174 ( teljes szöveg PDF -ként ).
- ↑ SIB: Fuselloviridae. Be: viruszone.expasy.org .
- ↑ NCBI: Halovírus VNH-1 (faj)
- ↑ Carolina M. Mizuno, Bina Prajapati, Soizick Lucas - Staat, Telesphore Sime - Ngando, Patrick Forterre, Dennis H. Bamford, David Prangishvili, Mart Krupovic, Hanna M. Oksanen: A Retba -tó új haloarchealis vírusai, amelyek megfertőzik a Haloferax és a Halorubrum fajokat. In: Környezeti mikrobiológia. 21. kötet, 6. szám, sfam, 2019. március 28., doi: 10.1111 / 1462-2920.14604 .
- ↑ NCBI: Halo vírus , Halo vírusok
- ↑ a b c Alon Philosof, Natalya Yutin, José Flores-Uribe, Itai Sharon, Eugene V. Koonin, Oded Béjà: Novel Abundant Oceanic Viruses of Uncultured Marine Group II Euryarchaeota. In: Aktuális biológia. 27. kötet, 9. szám), 2017. május 8., 1362-1368. O., Doi: 10.1016 / j.cub.2017.03.052 , PMC 5434244 (ingyenes teljes szöveg), PMID 28457865 .
- ↑ Luis H. Orellana, T. Ben Francis, Karen Krüger, Hanno Teeling, Marie-Caroline Müller, Bernhard M. Fuchs, Konstantinos T. Konstantinidis, Rudolf I. Amann: Niche differenciálódás az évente ismétlődő tengerparti tengerészcsoport II. Euryarchaeota között. In: Nature ISME Journal. 13. kötet, 2019. augusztus 26., 3014-3036. O., Doi: 10.1038 / s41396-019-0491-z .
- ↑ Xiaomin Xia, Wang Guo, Hongbin Liu: Basin Scale Variation on the Composition and Diversity of Archaea in the Pacific Ocean. In: Határok a mikrobiológiában. 2017. október 23., doi: 10.3389 / fmicb.2017.02057 .
- ↑ Ana-Belen Martin-Cuadrado et al. : A tengeri Euryarchaeota II csoport új osztálya a mediterrán mély klorofill maximumból. In: Nature ISME Journal. 2015. év 9. kötet, 1619–1634. O., Doi: 10.1038 / ismej.2014.249 .
- ↑ Daria Vaisman: Egyél meg . On: slate.com 2006 májusától.
- ↑ Bettina Hofer: Védekezés vírusokkal. Heise Technology Review, 2013. február 28., hozzáférés: 2014. augusztus 7 .
- ↑ JW Kretzer, R. Lehmann és mtsai: Bakteriofág endolizinek nagy affinitású sejtfal-kötő doménjeinek használata a bakteriális sejtek immobilizálásához és elválasztásához. In: Alkalmazott és környezeti mikrobiológia. 73. kötet, 2007, 1992-2000.
- ↑ C. Rozand, PCH Feng: Egy új, fágból származó ligandum specificitásának elemzése enzimhez kötött fluoreszcens vizsgálatban, Escherichia coli O157: H7 kimutatására. J. In: élelmiszervédelem. 72. kötet, 2009, 1078-1081.
- ↑ Bacteriophages - Új alkalmazások Food Microbiology ( Memento a március 2, 2013 az Internet Archive ) bioFood n ° 3, 2006. december 2. o..
- ↑ A fágligand -technológia alkalmazása (endotoxin eltávolítás, endotoxin kimutatás, élelmiszer -minőségi vizsgálat). Be: hyglos.de .
- ↑ DM Guglielmotti, DJ Mercanti, JA Reinheimer, ADL Quiberoni: A fizikai és kémiai kezelések hatékonysága a tej bakteriofágok inaktiválásában. In: Határok a mikrobiológiában. 2. kötet, 2012, doi: 10.3389 / fmicb.2011.00282 .
- ↑ Renata Filipa Cruz de Matos: Enterococcus faecalis V583 prophages: Dynamic kölcsönhatások és hozzájárulás a bakteriális patogén tulajdonságokhoz , értekezés, Universidade Nova de Lisboa (UNL), 2013. július
- ↑ NCBI: Phage P4 műhold (rang nélkül)
- ↑ SIB: 20 (Fuselloviridae) . Be: viruszone.expasy.org .
- ↑ SIB: Salterprovirus. 190 Salter provirus. , Itt: viruszone.expasy.org .
- ↑ NCBI: His 1 vírus (faj)
- ↑ SIB: Bicaudaviridae. Be: viruszone.expasy.org .
- ↑ ICTV: ICTV taxonómia története: Nitmarvirus NSV1 , EC 51, Berlin, Németország, 2019. július; E -mail megerősítés 2020. március (35. számú MSL)
- ↑ SIB: 113 (Inoviridae) on: viruszone.expasy.org .
- ↑ Stephen Nayfach, Simon Roux, Emiley A. Eloe-Fadrosh et al. : A Föld mikrobiómáinak genomi katalógusa. In: Természet . Biotechnológia, 2020. november 6., doi: 10.1038 / s41587-020-0718-6 , itt: S13 B. ábra
- ↑ ICTV: ICTV taxonómia története: Gammapleolipovirus His2 , EC 51, Berlin, Németország, 2019. július; E -mail megerősítés 2020. március (35. számú MSL)
- ↑ S. McGrath, D. van Sinderen D (szerk.): Bakteriofág: Genetics and Molecular Biology , 1.. Kiadás, Caister Academic Press, 2007, ISBN 978-1-904455-14-1 .
- ↑ izometrikus a vírusrészecskék esetében : megközelítőleg azonos térbeli expanzió minden irányban, például gömb vagy ikozaéder .
- ↑ SIB: Ampullaviridae. Be: viruszone.expasy.org .
- ↑ SIB: Bicaudaviridae. Be: viruszone.expasy.org .
- ↑ Elina Laanto, Sari Mäntynen, Luigi De Colibus, Jenni Marjakangas, Ashley Gillum, David I. Stuart, Janne J. Ravantti, Juha Huiskonen, Lotta-Riina Sundberg: A borealis tóban talált vírus összekapcsolja az ssDNA és a dsDNA vírusokat . In: Proceedings of the National Academy of Sciences. 114. kötet, 31. szám, 2017. július, doi: 10.1073 / pnas.1703834114 .
- ↑ SIB: Fuselloviridae. Be: viruszone.expasy.org .
- ↑ SIB: Globuloviridae. Be: viruszone.expasy.org .
- ↑ S. R. Krishnamurthy, D. Wang: A prokarióta riboszómális kötőhelyek kiterjedt megőrzése ismert és újszerű pikobirnavírusokban . In: Virológia . 516. kötet, 2018., 108-114. doi : 10.1016 / j.virol.2018.01.006 . PMID 29346073 .